植物の「冬支度」の新たなメカニズムを発見-細胞壁多糖β-1,4-ガラクタンが持つ機能-(大学院理工学研究科 高橋大輔助教)
2024/2/9
1 ポイント
●植物は気温の低下を感知すると凍結耐性を高めることができます。これを低温馴化(ていおんじゅんか)といい、植物が冬を越すための重要な生存戦略と考えられています。
●低温馴化では様々な変化が細胞内で起きていることが明らかになっていますが、植物細胞を取り囲んでいる「細胞壁」でどのような変化が起こっているのか、ほとんど明らかになっていませんでした。
●本研究により、細胞壁多糖の一種である「β-1,4-ガラクタン」が低温馴化により増加し、凍結耐性の上昇に寄与していることを発見しました。
2 概要
埼玉大学大学院理工学研究科の高橋大輔助教らと、大阪公立大学大学院理学研究科の曽我康一教授、東京工業大学生命理工学院の城所聡助教は、オーストラリアLa Trobe大学やドイツMax-Planck-Institute of Molecular Plant Physiologyの研究者らと共同で、植物の細胞壁1多糖の一種である「β-1,4-ガラクタン」が低温馴化により増加し、凍結耐性の上昇に寄与していることを発見しました。
本研究グループは、植物が気温の低下を感じて凍結環境への耐性を高める「冬支度」である低温馴化の過程(図1)において、細胞壁成分の一種であるガラクタンが増加することを突き止めました。このような反応は、本研究で主に用いたモデル植物シロイヌナズナだけではなく、ホウレンソウやシュンギクなどの他の凍結耐性が高い野菜でも広くみられる現象でした。また、β-1,4-ガラクタンは低温馴化過程での凍結耐性の向上や、組織の物理的性質の変化に寄与していることが明らかになりました。

シロイヌナズナは、低温(4℃)を予め処理することにより-10°Cでも生存することができる。
本成果は2024年2月8日 (アメリカ東部時間)に生物学全般を扱う米国の学術誌「Current Biology」に公開されました。
3 研究の背景
寒冷な環境に生育している植物は、厳しい凍結環境を生き抜くために、気温の低下を感知してより一層凍結耐性を高める「低温馴化」を行います(図1)。これは、植物にとって冬を越す前の「冬支度」だと言えます。これまでには低温馴化過程で、低分子の糖(グルコースやスクロースなど)の蓄積や細胞膜の脂質組成の変化、凍結耐性タンパク質の蓄積などが起こることが知られていましたが、細胞壁の変化はあまり調べられてきませんでした。
細胞壁は植物細胞の細胞膜の外側にあり、様々な多糖から成ります。植物の組織が凍結する際には、細胞と細胞の間で氷が形成されることがわかっており、細胞壁の重要性は以前から指摘されてきました。しかし、低温馴化過程でどのように変化し、それがどのような意味を持っているのかはわかっていませんでした。
4 研究内容
細胞壁はペクチン、ヘミセルロース、セルロースの三つに大別され、それぞれ様々な単糖が鎖状になった多糖とよばれる構造をとります。我々はまず、低温馴化することができて比較的凍結耐性の高い野菜(エンドウ、ホウレンソウ、コマツナ、シュンギク)を集め、破砕した組織から細胞壁を抽出し、多糖を分解して単糖の組成を分析することから始めました。その結果、細胞壁の主要な多糖の一つであるペクチンの中でガラクトースと呼ばれる単糖の割合が、これらの植物において低温馴化後に増加していることが明らかになりました。このガラクトースは、ペクチンの側鎖に見られるβ-1,4-ガラクタンと呼ばれる構造に由来すると考えられました(図2)。

植物細胞壁に含まれる多糖の一種であるペクチンには、側鎖としてガラクトース(黄色い丸)が連なったβ-1,4-ガラクタンがある。
このことを確かめるために、モデル植物の一つのシロイヌナズナ2を用いて様々なアプローチからβ-1,4-ガラクタンを解析しました。β-1,4-ガラクタンを特異的に認識する抗体3で組織中のβ-1,4-ガラクタンを可視化したところ、低温馴化処理によって組織全体でβ-1,4-ガラクタンが蓄積することがわかりました(図3)。他にも、β-1,4-ガラクタンを特異的に分解する酵素を用いた解析や、ガスクロマトグラフ質量分析計を用いた糖鎖結合分析4などから、同様のことが裏付けられました。
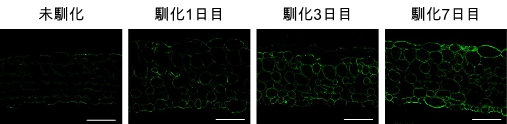
シロイヌナズナの葉において、低温馴化過程でβ-1,4-ガラクタン(緑色の蛍光)が組織全体に蓄積する様子が観察された。スケールバーは50 µmを表す。
β-1,4-ガラクタンは、生体内でGALSと呼ばれる酵素によってUDP-Galと呼ばれる材料物質から合成され、さらにUDP-GalはUGEと呼ばれる酵素によってUDP-Glcから変換されます。β-1,4-ガラクタンの合成を担うGALSやUGEといった遺伝子の発現量を解析したところ、低温馴化によって速やかに遺伝子発現量が上昇していることがわかりました。このことから、低温馴化過程におけるβ-1,4-ガラクタンの蓄積は、このような遺伝子の発現量調節によって起きていることが予想されます。
このβ-1,4-ガラクタンが蓄積する意味を探るために、β-1,4-ガラクタンの合成量が低下したシロイヌナズナ突然変異体5gals1 gals2 gals3を用いて、低温馴化後の凍結耐性を評価しました。その結果、野生型に比べて変異体では低温馴化後の凍結耐性が低下していることがわかりました(図4)。

低温馴化したシロイヌナズナを-10°Cの凍結温度に曝した後に、生きている組織が赤く染まる染色液に浸して凍結耐性を評価した。その結果、β-1,4-ガラクタンの合成量が低下したgals1 gals2 gals3は、野生型に比べて凍結耐性が低下していることがわかった。スケールバーは10 mmを表す。
また、引っ張り試験機を用いて低温馴化した葉の細胞壁の物理的強度を評価したところ、野生型の葉は低温馴化過程でかたくなっていくのに対し、gals1 gals2 gals3変異体ではそのような変化が見られませんでした。以上のことから、β-1,4-ガラクタンの蓄積は、低温馴化過程における凍結耐性の向上や組織の物性変化に貢献していることが示唆されました。
5 今後の展開
本研究では、上記の解析結果に加えて、低温馴化で誘導されるマスター転写制御因子であるDREB1/CBF6や、植物ホルモンであるアブシジン酸(ABA)7がUGEの遺伝子発現を制御している可能性も示唆されています(図5)。今後は、どのようなメカニズムでβ-1,4-ガラクタンが蓄積しているのかを明らかにするために、DREB1/CBFやABAとの関係にも着目して細胞壁関連遺伝子を解析していく必要があります。
また、β-1,4-ガラクタンによる組織の物性変化が、実際にどのようなメカニズムで凍結耐性の向上に貢献しているのか明らかにしていく必要があります(図5)。本研究で低温馴化によるβ-1,4-ガラクタンの蓄積は広範な植物で見られたため、これらのことが明らかになれば様々な植物での低温馴化過程での物性変化や凍結耐性の改変を行うことができ、農作物の収量増加や品質の向上にもつながる可能性があります。
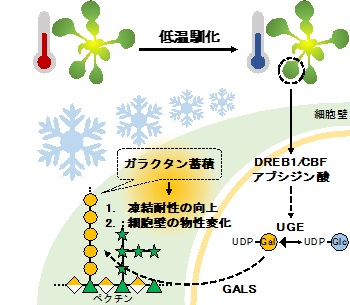
低温を感知すると、DREB1/CBFやABAの誘導によりUGEの発現が増加し、β-1,4-ガラクタン合成の材料となるUDP-Galの合成が促されると考えられる。その後、GALSによってβ-1,4-ガラクタンが合成され、凍結耐性の向上や細胞壁の物性変化を導くと予想される。今後は、低温馴化過程でのβ-1,4-ガラクタン合成の制御機構やβ-1,4-ガラクタンが蓄積する生物的意義をより詳細に明らかにする必要がある。
6 原論文情報
| 掲載誌 | Current Biology |
|---|---|
| 論文名 | Structural changes in cell wall pectic polymers contribute to freezing tolerance induced by cold acclimation in plants |
| 著者名 | Daisuke Takahashi, Kouichi Soga, Takuma Kikuchi, Tatsuya Kutsuno, Pengfei Hao, Kazuma Sasaki, Yui Nishiyama, Satoshi Kidokoro, Arun Sampathkumar, Antony Bacic, Kim L. Johnson, Toshihisa Kotake |
| DOI | 10.1016/j.cub.2024.01.045 |
| URL | https://doi.org/10.1016/j.cub.2024.01.045 |
7 研究支援
本研究は、日本学術振興会科研費(20 K15494、23K05144、16 K07391、18H05495、19 K06702)、市村清新技術財団植物研究助成(29-14、30-12)、加藤記念バイオサイエンス財団研究助成、日本科学協会笹川科学研究助成(2023-4014)、山下太郎顕彰育英会の支援を受けて行われました。
8 用語解説
1. 細胞壁
植物細胞が動物細胞と大きく異なる点が「細胞壁」があることです。細胞壁は植物細胞の一番外側にある多糖やフェノール性化合物が複雑に絡んだ構造を持つ部分で、力学的強度が高いことが特徴です。そのため、樹木などの植物がかたくて丈夫なのは細胞壁が豊富にあるからと言われています。また、地球上で最も多い炭素資源であると言われ、バイオエタノールの原料としても注目されています。
2. シロイヌナズナ
大腸菌やショウジョウバエなどと同様に、植物において用いられるモデル生物です。ゲノムサイズが小さく、生育が早いため実験で扱いやすいことが特徴です。
3. 抗体
特定の物質(タンパク質や糖鎖など)を認識して結合するタンパク質を抗体と呼びます。この抗体の認識特異性を利用して、植物組織上に存在する特定の多糖に抗体を結合させて標識することで、植物組織のどの部位に特定の多糖が蓄積しているのかを可視化することができます。これを免疫組織染色と言います(図3)。
4. 糖鎖結合分析
細胞壁多糖を構成する単糖は、それぞれ特有の結合様式によって互いにつながっています。この結合様式を定量的に分析する手法が糖鎖結合分析です。この分析にはガスクロマトグラフ質量分析計という装置を用います。
5 . 突然変異体
モデル植物シロイヌナズナでは、様々な遺伝子が機能不全になった突然変異体系統がカタログ化されています。本研究では、GALS1、GALS2、GALS3という三つの酵素が働かなくなった三重変異体を用いて解析を行いました(図4)。
6. DREB1/CBF
植物が低温に曝された時に誘導される転写因子の一種です。転写因子とは、生体内の様々な遺伝子の発現を制御しているタンパク質のことです。低温馴化過程では、DREB1/CBFの発現誘導が鍵となって他の多くの遺伝子の発現パターンを制御しています。
7. アブシジン酸(ABA)
植物の休眠や気孔の閉鎖などを制御する植物ホルモンです。低温や乾燥などの環境刺激に応答することで蓄積が促されますDREB1/CBFと同様、アブシジン酸も様々な遺伝子の発現を制御しています。
参考URL
