植物は光環境を認識して光合成の電子受容体量を調節する ~使わない電子受容体を貯めない仕組みを発見~(大学院理工学研究科 川合真紀教授 共同研究)
2023/11/13
概要
一般財団法人電力中央研究所(電中研、理事長:平岩芳朗)サステナブルシステム研究本部生物・環境化学研究部門の橋田慎之介上席研究員、国立大学法人埼玉大学(埼玉大、学長:坂井貴文)大学院理工学研究科の川合真紀教授、株式会社セレス(セレス、代表取締役社長:松村卓郎)技術本部環境部の福田裕介、石山知波は共同で、光合成の電子受容体量を調節する仕組みを世界に先駆けて明らかにしました。光エネルギーを化学エネルギーに変換する光合成は、植物の成長の根幹となる現象です。光エネルギーによって葉緑素(クロロフィル)から取り出された電子は受容体(NADP+)に受け渡されてNADPHという物質で一時的に保存されます。この受容体量が光環境によって変化する現象は1959年に発見されましたが、その生理現象の全体像はいまだに解明されていません。
本研究では受容体の精密定量技術を駆使した研究を進め、光が当たる昼間の葉緑体内部で受容体の合成が促進される現象が、葉緑体ストロマ※1内のpH制御によって調節される仕組みを明らかにしました。
これまで、光合成活性のシミュレーション等では電子受容体量は一定と仮定されてきましたが、受容体量も光合成に大きく関与することが本研究によって初めて明らかになりました。電子受容体量を調節することで植物成長が制御される知見は、光合成出力の改善によるバイオマス増産のみならず有害植物の成長抑制に資する新たな農薬開発にもつながる重要な成果といえます。
本研究成果は、2023年11月6日付けで、英国科学雑誌のNature Communicationsに掲載されました。
1.背景
植物は太陽光を浴びて光合成を行い、光エネルギーを化学エネルギー(ATP)※2に変換して成長します(図1)。この過程では、葉緑体から取り出された電子が電子伝達経路※3を通じて受容体(NADP+)に受け渡されてNADPH(還元力)に一時的に保存され、同時にATP(化学エネルギー)が生産されます。これらを利用してCO2を固定し、糖やデンプンを合成しています。光がない夜には電子伝達が起こらないため、NADP+の状態で蓄積していると考えられてきましたが、近年、我々のグループではNADP+の高精度定量技術を開発し、30分以上光が当たらない葉では、受容体を合成できない変異植物と同等レベルまで減少することが分かってきました。野外環境では太陽光は雲や障害物によって遮られます。これにより、実際に葉にあたる光の強さや時間は日中を通して大きく変動します。数分間の日陰では受容体は減少しませんが、長時間の日陰や夜には減少するため、植物が光環境を認知して受容体量を増減していると考えられます。しかし、その調節の仕組みは不明でした。
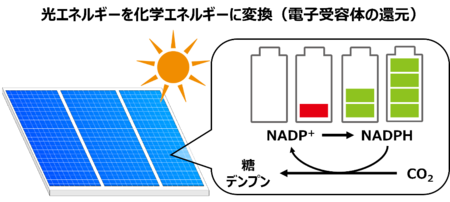
図1:光合成による光エネルギーの保存
光合成では葉に光が当たると葉緑体内で電子伝達経路が駆動し、受容体であるNADP+が還元されてNADPHが生産されます。このNADPHが還元力として使用されることでCO2を固定し、糖やデンプンが合成されます。
2.研究成果の特長
① 葉緑体内部の受容体量の変動と合成活性特性の計測
電子受容体であるNADP+は細胞に広く存在します。光合成以外の反応にも利用されるため、はじめに葉緑体特異的な受容体量の変動について調べました。植物の葉から単離した無傷の葉緑体を用いて受容体量を計測する実験系と受容体の合成活性を計測する実験系を開発し、光に応答した受容体の変動が葉緑体で起こることを証明しました。また、受容体の合成酵素の活性には光が必要であるとともに、アルカリ性のpHで活性が顕著に増大することを明らかにしました(図2)。
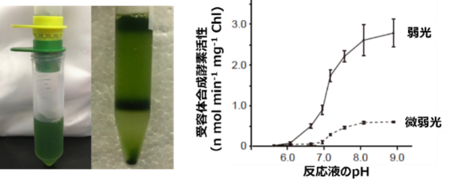
図2:無傷葉緑体単離の様子(左)と葉緑体破砕液に含まれる電子受容体合成活性のpH依存性(右)
葉緑体に照射する光強度に比例して合成活性が高まると共に、反応液のpHがアルカリ性の時に活性が増大するという結果が得られました。
② 電子伝達経路と受容体合成の関係を解明
光合成の電子伝達経路は、受容体がなければ電子が渋滞してしまい動きませんが、サイクリック電子伝達経路※4というもう一つの経路が同時に駆動しています。このサイクリック電子伝達経路では電子を受容体に渡すことなく系内を循環することで、葉緑体内のpH勾配※5を光がある時の状態に保ちATPを合成します。電子伝達経路の阻害剤によって全ての電子伝達を止めると、光があっても受容体は合成されません。ここで、サイクリック電子伝達経路を特異的に阻害した時にも受容体の合成が抑制されました。また、サイクリック電子伝達経路を欠損する変異体では受容体合成が顕著に遅延しており、光に応答した速やかな受容体の増加にはこの経路の駆動によるpH勾配が重要であることが明らかになりました。
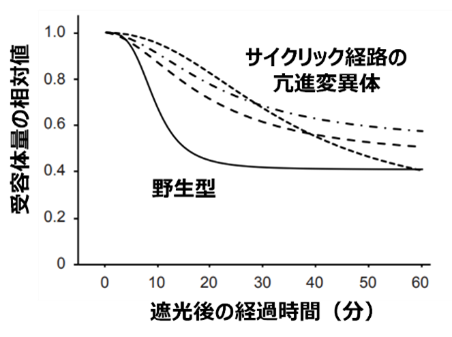
図3 サイクリック電子伝達活性と受容体現象の関係
③ サイクリック経路と受容体分解の関係を解明
植物の葉に光を当てた後に遮光すると、受容体量は徐々に減少します。この時、遮光前に強光を照射することでサイクリック経路を亢進すると受容体の減少は遅延しました。同様に、この経路の亢進変異体では受容体の減少が遅延し(図3)、欠損変異体では減少が加速しました。また、受容体の分解活性は弱酸性から中性のpH条件で活性化し、アルカリ条件では消失していました。従って、遮光によって生じる葉緑体内部のpH変化と受容体分解酵素の活性特性が、一致することが分かりました。さらに、遮光時に葉緑体内のpHを調節するイオン輸送体の欠損変異体でも同様の遅延が観察されたことから、葉緑体内部の受容体量が光環境に依存したpH変化によって調節されることがわかりました。
以上の成果をまとめると、植物の葉の葉緑体では、光によって電子伝達経路が駆動することで葉緑体ストロマのpHが受容体を合成しやすい環境に調整され、受容体量が増加して光合成出力が増大すると考えられます(図4)。光が遮られた環境ではサイクリック電子伝達経路によってpHが維持され、数分間は次の光まで待機しますが、循環可能な電子が尽きると、葉緑体ストロマのpHは受容体を分解しやすい環境に調整されて減少することが分かってきました。
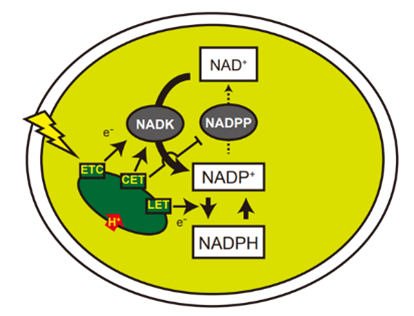
図4 本研究の成果から考えられる光照射時の受容体量調節の仕組み
光によって電子伝達経路(ETC)が駆動することで受容体合成酵素が活性化し(ETCからNADKへの矢印)、この時にサイクリック電子伝達経路(CET)の働きによってpHがアルカリ性に調節されることで受容体合成が促進され(CETからNADKへの矢印)、受容体分解が抑制されます(CETからNADPPへのTバー)。受容体が増加することで光エネルギーを化学エネルギーとして保存する経路(LET)が充分に駆動されるようになり光合成出力が高まります(LETからの矢印)。
3.本研究成果の意義と今後の展望
これまでに、光合成活性の改善や最適化のためのシミュレーションでは、電子受容体の変動についてはあまり考慮されてきませんでした。受容体の量は、実験室内の光環境によって容易に変動してしまうため、適切な評価が困難であったことが原因の一つでもありました。本研究から、変動する光環境に応答した受容体増減の仕組みが分かりました。光合成出力の改善によるバイオマス増産のみならず有害植物の成長抑制に資する新たな農薬開発にもつながる重要な発見といえます。また、これまでの教科書に載っていた受容体の記載では、常に一定量があることが前提とされたものでしたが、光合成活性の制御要因の一つとして認識を新たにするものでもあります。こうした受容体量を調節する仕組みに関与する酵素遺伝子など、未だ明らかになっていない構成要素を同定することは、新たな植物科学理論に基づく成長制御技術の開発に寄与すると考えられるため、さらに詳しく研究を進めてその機能を明らかにしていきます。
用語説明
※1 葉緑体ストロマ:葉緑体内部に分布する水性領域であり、様々な可溶性酵素タンパク質を含んでいる。
※2 化学エネルギー(ATP):生体内に広く分布する物質であり、広範な物質の生合成反応や代謝反応において重要な役割を担う。
※3 電子伝達経路:リニア電子伝達経路とも呼ばれ、ATPやNADPHを生産することで、その後のCO2固定を始めとしたさまざまな代謝経路にエネルギーと還元力を供給する役割を担う。
※4 サイクリック電子伝達経路:光合成の電子伝達経路の1つで、電子が受容体のNADP+に受け渡されずにNADPHが生産されない経路とされる。電子が系内を循環することで葉緑体内のpH勾配が維持され、ATPのみが生産される。
※5 葉緑体内のpH勾配:光によって電子伝達経路が駆動すると、ストロマ側のプロトンがチラコイド膜内に輸送されることで、チラコイド内のpHは酸性側に傾き、ストロマ内のpHはアルカリ化に傾くpH勾配が形成されます。
論文タイトルと著書
| 論文タイトル | Adjustment of light-responsive NADP dynamics in chloroplasts by stromal pH |
|---|---|
| 著者 | Yusuke Fukuda (セレス), Chinami Ishiyama (セレス), Maki Kawai-Yamada (埼玉大), Shin-nosuke Hashida (電中研) |
| 掲載誌 | Nature Communications |
| DOI | 10.1038/s41467-023-42995-9 |
なお、本研究はJSPS 科研費 基盤研究20K06695、新学術領域研究17H05714、新学術領域研究19H04715の助成を受けたものです。
参考URL
