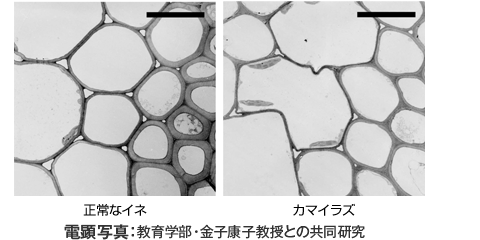
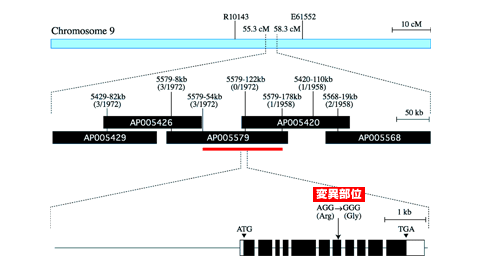
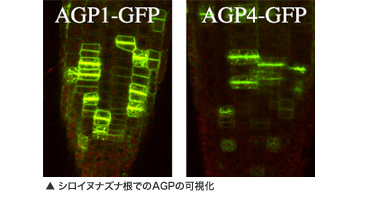
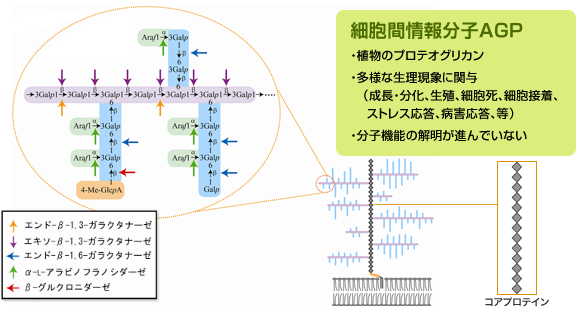
TOP
ごあいさつ
掲載者一覧